Transmission des caractères
Exemple de transmission aléatoire de deux caractères au cours de la genèse des spermatozoïdes (méiose)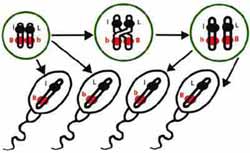
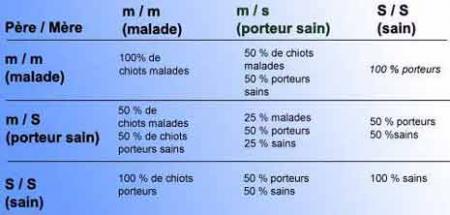
Prévision statistique sur une portée issue d’un accouplement entre deux parents double hétérozigotes pour deux caractères
Lors de la formation des cellules sexuelles (gamétogénèse), un processus complexe appelé "méïose" dissocie les 39 paires de chromosomes, les mélange et redistribue comme un jeu de cartes une combinaison de 39 chromosomes à chacun des gamètes. La diversité génétique des gamètes assure ainsi la variabilité génétique au sein de chaque race canine. L'union d'un spermatozoïde et d'un ovule (fécondation) aboutit à la formation de l'oeuf (zygote) dans lequel les chromosomes hérités des deux parents se réunissent à nouveau par paires homologues.
On voit donc qu'une sélection involontaire s'opère déjà à deux niveaux :- lors de la méïose où une information génétique différente est distribuée à chaque gamète,- lors de la fécondation puisqu'on ne peut prédire quels spermatozoïdes seront fécondants ni quels ovules seront fécondés.
Ajoutons à cela les possibilités de mutation des gènes (chaque individu a, au moment de sa conception, environ une chance sur 10 pour qu'un de ses gènes ait muté) et nous comprenons alors pourquoi il est possible d'orienter sa sélection en élevage mais certainement pas de la maîtriser complètement ! ...
Naturellement, chaque chromosome ne porte pas qu'un seul gène. Chaque locus correspond à une information génétique qui peut aussi bien coder pour la longueur du poil que pour l'intensité de sa couleur ou pour une aptitude enzymatique ou encore moduler l'expression d'un autre gène (épistasie).
Certains caractères sont sous la dépendance de plusieurs gènes et la dominance d'un allèle sur son homologue peut être incomplète.
Enfin, notons que nous sommes très loin de maîtriser l'ensemble des combinaisons génétiques, ce qui inspire l'humilité et la prudence dans ce domaine soumis aux lois des probabilités.
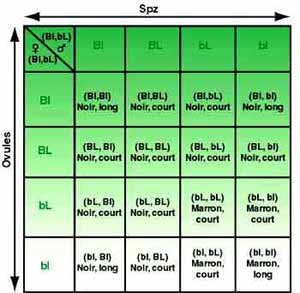
Cependant, l'éleveur peut déjà s'exercer pour deux caractères (longueur et couleur du poil), à prévoir le phénotype des chiens ayant hérité des combinaisons suivantes, sachant que pour le gène "longueur du poil", l'allèle L (poils courts) domine l'allèle "l" (poils longs)
- BL / BL : poils courts, robe noire, individu double homozygote ;- Bl / bL : double hétérozygote à poils courts et robe noire car "B" et "L" dominent "b" et "l" ;- bl / bl : double homozygote à poils longs et bruns. C'est la seule combinaison permettant aux deux gènes récessifs de s'exprimer car ils sont chacun en double exemplaire ;- bL / bl : Homozygote pour "b", hétérozygote pour la longueur du poil. Ce chien aura le poil court et brun ;- Bl / bl : Homozygote pour "l", hétérozygote pour la couleur. Individu à poil long et noir.
Ces 5 exemples (parmi les 16 combinaisons possibles) permettent déjà à l'éleveur de se familiariser avec le raisonnement génétique.
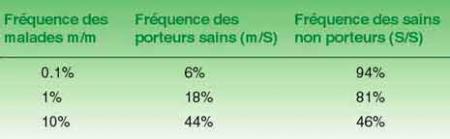
Ainsi, pour reprendre l'exemple précédent, si l'on ne s'intéresse qu'aux deux caractères étudiés, les "chances" de trouver parmi la population, un chien marron à poils longs sont estimées à 6,25% (1/16ème), 18,75% (3/16èmes) pour les poils longs et noirs ou courts et marron et 56,25% (9/16 èmes) pour les poils courts et noirs (voir tableau).
Connaissant le caractère dominant ou récessif d'une qualité ou d'une tare, l'éleveur peut ainsi orienter sa sélection pour mettre toutes les chances de son côté ...
L'apparence est-elle le reflet du programme ?
Nous avons vu qu'il existait des caractères dominants et des caractères récessifs. Pour reprendre l'exemple précédent concernant la couleur de la robe, le Noir (B) domine le marron (b). Lorsque l'on voit un chien marron, il est facile de deviner son génotype qui ne peut être que b/b puisque le caractère récessif marron ne peut s'exprimer qu'à l'état homozygote. Dans le cas des caractères récessifs, le phénotype est donc un bon reflet du génotype. En revanche, le phénotype "noir" peut correspondre soit au génotype B/b (hétérozygote), soit au génotype B/B (homozygote), les gènes ayant toujours au moins deux possibilités de s'exprimer.
- dans le premier cas, le chien est noir mais porteur d'un allèle marron qu'il pourra transmettre à sa descendance.
- dans le deuxième cas, le chien homozygote transmettra obligatoirement un de ses deux allèles B à sa descendance qui sera donc uniformément noire en première génération (appelée F1) quels que soient les allèles apportés par son partenaire.
L'aspect d'un chien ne reflète donc pas isolément son patrimoine génétique.
Le principe même de la confirmation qui donne au candidat l'autorisation officielle de reproduire en évaluant son génotype en fonction de son phénotype, est donc sujet à controverse.
Il est cependant possible de se faire une idée du génotype d'un étalon par son aptitude à marquer sa descendance pour le caractère considéré.
En effet, si un chien ne compte que des chiots noirs parmi sa descendance lorsqu'il est accouplé avec une chienne marron (donc obligatoirement b/b), il devient très probable qu'il soit homozygote B/B. Dans ce cas, tous les produits sont B/b, donc noirs.
S'il était B/b, il aurait 50% de chances de donner naissance à des chiots marrons à l'issue de ce même accouplement et il serait donc exceptionnel de ne pas en trouver parmi sa descendance. L'effectif de la portée permet donc également de renforcer l'estimation génétique d'un reproducteur.
Une portée d'un ou deux chiots reste évidemment peu significative.
On comprend dès lors que si l'éleveur désire fixer un caractère qu'il juge positif pour son élevage, il devra rechercher des reproducteurs qui présentent tous deux, non seulement ce caractère mais aussi un lien de parenté (consanguinité) de manière à augmenter les chances d'homozygotie, c'est-à-dire l'uniformité des ordres édictés par les gènes parentaux.
A titre d'exemple, si un éleveur désire fixer le caractère marron, il n'aura qu'à accoupler des reproducteurs marrons.
S'il considère la robe marron comme un défaut, il lui faudra alors sélectionner des reproducteurs noirs qui, en outre, ne comptent aucun chien marron parmi leur généalogie.
On devrait donc se réserver la possibilité d'annuler une confirmation au cas où un défaut héréditaire apparaîtrait dans la descendance d'un reproducteur ...